
What is cadherin?
The cadherin glycoproteins includes over 100 members divided into 6 subfamilies; type I classical cadherins , type II atypical cadherins, desmosomal cadherins, flamingo cadherins, proto-cadherins and several ungrouped members. Cadherins can be identified through common motifs in their extracellular domains termed cadherin repeats. Not all cadherins are involved in cell-cell adhesion, though type I and type II cadherins have well established roles in this process [1]. Both of these subfamilies contain cadherin repeats within their extracellular domains, with the outermost cadherin repeats facilitating extracellular interactions with cadherins on apposing cells (transinteractions). Type I cadherins can in addition engage in lateral interactions on the same cell (cis interactions)[2]. Intercellular interactions between cadherins can occur between those of the same type (homophilic binding) or a different type (heterophilic binding). Intracellular interactions involve the cytoplasmic domains of the cadherins. In the case of type I cadherins these interactions can be used to identify this subfamily, namely through their ability to bind catenins via their cytoplasmic tails. Catenins form part of the bridge connecting adherens junctions to the actin cytoskeleton. It should be noted that individual cadherin interactions are weak. The strength of cadherin-based adhesive junctions comes from the clustering of multiple, weak cadherin-cadherin interactions [3].
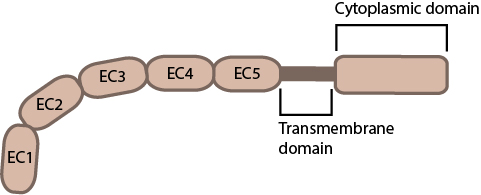
This schematic diagram illustrates the molecular organization of cadherin.
The cadherin protein family are common cell-adhesion molecules (CAMs) that mediate cell-cell contacts at anchoring junctions (e.g. adherens junctions, desmosomes) and at prominent sites of cell-cell communication (e.g. neuronal synapses). There are over 100 different cadherin family members that are grouped into at least 6 subfamilies, including type I classical cadherins, type II atypical cadherins and desmosomal cadherins [4]. All cadherins share a common architecture in their extracellular domain that comprises cadherin repeats, with classical cadherins containing five of these repeats. Subtle differences between cadherins impart each type with specificity for particular tissue and cell types. Cadherins use a common set of adaptor molecules and pathways to facilitate cell adhesion and communication, however the strength of adhesion varies with the type of cadherin present [5].
Cadherin interactions during adherens junction formation
Most cadherins adhere by homophilic interactions (i.e. they bind to the same type of cadherin) but certain types (e.g. E-cadherin) also adhere by heterophilic interactions (i.e. they bind other types of cadherin). Cadherin association is sensitive to extracellular calcium (hence their name, calcium adhering). The interactions can take place laterally on the same cell, called a cis interaction, or between two cells, called a trans interaction.
Structurally, classical cadherins have five Ca+2-dependent extracellular domains and a relatively short cytoplasmic domain. Although cadherin-cadherin binding between the extracellular domains is relatively weak, the conformational changes that are induced after binding imparts the individual cadherins with rigidity. This stabilizes the interaction and fosters additional lateral cis interactions with other cadherins and generates tighter adhesions. Increased clustering of cadherins at sites of cell-cell contact correlates with increased stability and maturation of actin-based structures such as dendritic spines [6].
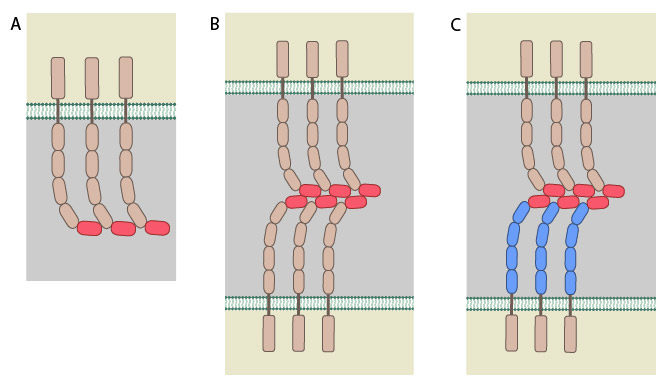
All cadherins have a common extracellular domain that is structured into tandem blocks that are variable in number and are called ‘cadherin repeats’; classical cadherins have five cadherin repeats (as shown). The cadherin intracellular domain is bound by adaptor proteins. (A) Example of a cis interaction (i.e. on the same cell) that is homophilic (i.e. cadherins of the same type). (B) Example of a trans interaction (i.e. between two cells) that is homophilic. (C) Example of a trans interaction that is heterophilic (between two different types of cadherin).
The classical cadherins (e.g. E-, N-, and P-cadherins) are the most common family members. Classical cadherins interact directly with p120ctn at their transmembrane region and through their cytoplasmic tails to beta (β)-catenin or plakoglobin(i.e. gamma [γ]-catenin). The correct function and stability of the cadherins requires these associations (reviewed in [7]). β-catenin binds tightly to classical cadherins before they are transported to the cell surface [8][9]). Cadherins further interact indirectly with other adaptor proteins (e.g. alpha [α]-catenin, vinculin, EPLIN,α-actinin, zyxin) to form linkages between the cell membrane and the actin cytoskeleton (reviewed in [10]). Desmosomes in contrast, have two specialized cadherins that interact with specific adaptor proteins (e.g. plakoglobin, plakophilin, desmoplakin) to form links with the intermediate filaments.
Cadherin subfamilies:
- Type I classical cadherins – includes epithelial (E)-, neural (N)- and placental (P)-cadherin
- Type II atypical cadherins – includes vascular endothelial (VE)-cadherin
- Desmosomal cadherins – includes desmoglein and desmocollin
- Flamingo cadherins
- Proto-cadherins
- Ungrouped cadherins
Non-junctional cadherin clusters
In addition to the well-established role of classical cadherins in mediating intercellular adhesions, recent research has also revealed the existence of distinct cadherin clusters outside of these adhesions. The physiological significance of such non-junctional cadherin clusters was elucidated in a recent study that analyzed the organization and functions of HMR-1, the C.elegans homolog of cadherins, within its single-celled zygote that is devoid of any intercellular junctions. The study demonstrated the organization of HMR-1 into clusters and the ability of these clusters to associate with cortical actin during the retrograde flow of the cortex. Surprisingly, HMR-1 clusters were shown to negatively regulate myosin-mediated contractility of the actin cortex and, as a result, impede its retrograde flow by inhibiting Rho-I GTPase functions and the subsequent accumulation of non-muscle myosin II (NMY-2) at the cortex. An increase in the levels of HMR-1 was also associated with a decline in the initiation and ingression rate of the cytokinetic furrow during the first division of the zygote .The findings from the study confirm the existence of adhesion-independent roles for HMR-1 clusters in reinforcing the stability of the actomyosin cortex and regulating cell-shape dynamics during the very early phase of embryo development [27989674].
References
- Niessen CM, Leckband D, and Yap AS. Tissue organization by cadherin adhesion molecules: dynamic molecular and cellular mechanisms of morphogenetic regulation. Physiol Rev 2011; 91(2):691-731. [PMID: 21527735]
- Harrison OJ, Jin X, Hong S, Bahna F, Ahlsen G, Brasch J, Wu Y, Vendome J, Felsovalyi K, Hampton CM, Troyanovsky RB, Ben-Shaul A, Frank J, Troyanovsky SM, Shapiro L, and Honig B. The extracellular architecture of adherens junctions revealed by crystal structures of type I cadherins. Structure 2011; 19(2):244-56. [PMID: 21300292]
- Baumgartner W, Hinterdorfer P, Ness W, Raab A, Vestweber D, Schindler H, and Drenckhahn D. Cadherin interaction probed by atomic force microscopy. Proc. Natl. Acad. Sci. U.S.A. 2000; 97(8):4005-10. [PMID: 10759550]
- Hill E, Broadbent ID, Chothia C, and Pettitt J. Cadherin superfamily proteins in Caenorhabditis elegans and Drosophila melanogaster. J. Mol. Biol. 2001; 305(5):1011-24. [PMID: 11162110]
- Chu Y, Eder O, Thomas WA, Simcha I, Pincet F, Ben-Ze’ev A, Perez E, Thiery JP, and Dufour S. Prototypical type I E-cadherin and type II cadherin-7 mediate very distinct adhesiveness through their extracellular domains. J. Biol. Chem. 2005; 281(5):2901-10. [PMID: 16253998]
- Johnson CP, Tang H, Carag C, Speicher DW, and Discher DE. Forced unfolding of proteins within cells. Science 2007; 317(5838):663-6. [PMID: 17673662]
- Stemmler MP. Cadherins in development and cancer. Mol Biosyst 2008; 4(8):835-50. [PMID: 18633485]
- Aberle H, Butz S, Stappert J, Weissig H, Kemler R, and Hoschuetzky H. Assembly of the cadherin-catenin complex in vitro with recombinant proteins. J. Cell. Sci. 1994; 107 ( Pt 12):3655-63. [PMID: 7706414]
- Hinck L, Näthke IS, Papkoff J, and Nelson WJ. Dynamics of cadherin/catenin complex formation: novel protein interactions and pathways of complex assembly. J. Cell Biol. 1994; 125(6):1327-40. [PMID: 8207061]
- Parker KK, Brock AL, Brangwynne C, Mannix RJ, Wang N, Ostuni E, Geisse NA, Adams JC, Whitesides GM, and Ingber DE. Directional control of lamellipodia extension by constraining cell shape and orienting cell tractional forces. FASEB J. 2002; 16(10):1195-204. [PMID: 12153987]