
What are stress fibers?
Stress fibers are higher order cytoskeletal structures composed of cross-linked actin filament bundles, and in many cases, myosin motor proteins, that span a length of 1-2 micrometers [1]. At least 4 types of stress fibers have been identified in mammalian cells [2]. These are dorsal stress fibers, ventral stress fibers, transverse arcs, and the recently identified perinuclear actin cap, which is an important mediator in nuclear mechanotransduction. Each type of stress fiber is defined by their location in the cell, their morphology and their function at focal adhesions (reviewed in [3]). The presence of motor proteins in stress fibers enables contractility – an important factor in stress fiber function and in cell motility. In most cases, stress fibers connect to focal adhesions, and hence are crucial in mechanostransduction. In mammalian cells, stress fibers undergo continuous assembly and disassembly. This allows them to maintain cellular tension and undergo modification in response to various forces (e.g., mechanical stress [4][5]).
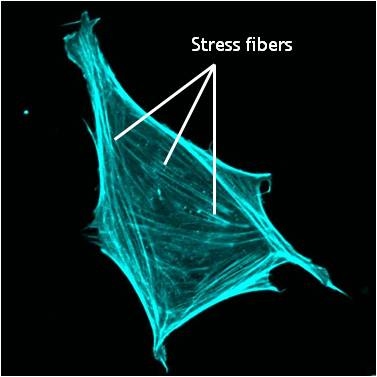
A mouse embryonic fibroblast of the RPTPa cell line, plated on a fibronectin coated glass cover slip. The cell was transfected with RFP-Lifeact (a kind gift from Dr Roland Wedlich-Soldner, Max Planck Institute of Biochemistry, Germany), which labels F-actin in living cells. The cell was imaged using a Nikon A1Rsi confocal microscope at 60x magnification and false coloured cyan. Image captured by Wei Wei Luo, Mechanobiology Institute, Singapore.
Stress fiber assembly can be described in a series of defined steps. Here, ~10-30 parallel actin filaments are bundled together via cross-linking proteins. α-actinin; for example,forms bands along the stress fibers [6] while filamin [7], binds uniformly along the length of the fibers [8]. In ventral stress fibers and transverse arcs, bands of α-actinin appear to alternate with bundles of non-muscle myosins (e.g. myosin II) at a periodicity of ~1-2 µm [8][9]. Although the contractile nature of transverse arcs and ventral stress fibers resembles the contraction of muscle sarcomeres, the organization of the actin filaments, actin binding proteins, and myosin bundles is considerably more random in stress fibers (when compared to muscle fibers) and the polarity of the actin filaments does not strictly alternate along the bundles [1][10][11][12] (reviewed in [13][3][14]). Besides myosin and α-actinin, stress fibers also contain many other proteins like fascin [15], caldesmon and tropomyosin [16], and regulatory proteins like myosin light chain kinase [12]. The exact functions of the majority of these proteins in stress fiber activity and assembly are still largely unclear.
Types of Stress Fibers
Transverse arcs (TAs) are found only in cells that are actively protrusive. TA appear as curved bundles of actin and myosin filaments behind the lamella. Unlike the ventral fibers, TAs do not interact directly with focal adhesions (FA). TAs have been implicated in actin retrograde flow from the leading edge to the cell center where they are disassembled [17].
Dorsal stress fibers (DSF) are the main transmitters of contractile forces (as opposed to producers) to the underlying substrate. They attach to FAs at the base of the cell then rise towards the dorsal surface to form a loose matrix of actin filaments [18]. DSF often terminate at a TA at their proximal end. It is important to note that DSF are suggested to serve as precursors of VSF and do not exhibit periodic distribution of α-actinin and myosin on the filaments (aka contractile bundle), so their identity as stress fibers has been debated (reviewed in [3]).
Ventral stress fibers (VSF) are filament bundles located at the ventral surface of the cell that are attached to FAs at each end of the bundle; these stress fibers extend from FAs close to the cell edge (i.e. lamellipodia) to an adhesion behind or near the nucleus (reviewed in [19]). The VSF are fundamental to tail retraction and cell shape changes during cell migration [20] and on their sides they can structure cell borders against inward pressure of the membrane [21].
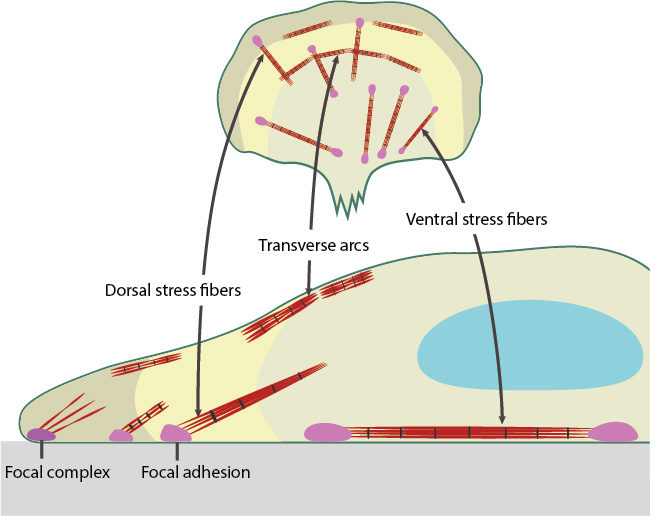
Dorsal and Ventral stress fibers connect to focal adhesion and transmit contractile force. Transverse arc stress fibers drive retrograde actin flow in actively protruding cells.
References
- Cramer LP, Siebert M, and Mitchison TJ. Identification of novel graded polarity actin filament bundles in locomoting heart fibroblasts: implications for the generation of motile force. J. Cell Biol. 1997; 136(6):1287-305. [PMID: 9087444]
- Tojkander S, Gateva G, and Lappalainen P. Actin stress fibers–assembly, dynamics and biological roles. J Cell Sci 2012; 125(Pt 8):1855-64. [PMID: 22544950]
- Naumanen P, Lappalainen P, and Hotulainen P. Mechanisms of actin stress fibre assembly. J Microsc 2008; 231(3):446-54. [PMID: 18755000]
- Hirata H, Tatsumi H, and Sokabe M. Dynamics of actin filaments during tension-dependent formation of actin bundles. Biochim. Biophys. Acta 2007; 1770(8):1115-27. [PMID: 17498881]
- Kaunas R, Nguyen P, Usami S, and Chien S. Cooperative effects of Rho and mechanical stretch on stress fiber organization. Proc. Natl. Acad. Sci. U.S.A. 2005; 102(44):15895-900. [PMID: 16247009]
- Lazarides E, and Burridge K. Alpha-actinin: immunofluorescent localization of a muscle structural protein in nonmuscle cells. Cell 1975; 6(3):289-98. [PMID: 802682]
- Wang K, Ash JF, and Singer SJ. Filamin, a new high-molecular-weight protein found in smooth muscle and non-muscle cells. Proc. Natl. Acad. Sci. U.S.A. 1975; 72(11):4483-6. [PMID: 53835]
- Pavalko FM, Otey CA, and Burridge K. Identification of a filamin isoform enriched at the ends of stress fibers in chicken embryo fibroblasts. J. Cell. Sci. 1989; 94 ( Pt 1):109-18. [PMID: 2693470]
- Weber K, and Groeschel-Stewart U. Antibody to myosin: the specific visualization of myosin-containing filaments in nonmuscle cells. Proc. Natl. Acad. Sci. U.S.A. 1974; 71(11):4561-4. [PMID: 4612524]
- Langanger G, Moeremans M, Daneels G, Sobieszek A, De Brabander M, and De Mey J. The molecular organization of myosin in stress fibers of cultured cells. J. Cell Biol. 1986; 102(1):200-9. [PMID: 3510218]
- Begg DA, Rodewald R, and Rebhun LI. The visualization of actin filament polarity in thin sections. Evidence for the uniform polarity of membrane-associated filaments. J. Cell Biol. 1978; 79(3):846-52. [PMID: 569662]
- Peterson LJ, Rajfur Z, Maddox AS, Freel CD, Chen Y, Edlund M, Otey C, and Burridge K. Simultaneous stretching and contraction of stress fibers in vivo. Mol. Biol. Cell 2004; 15(7):3497-508. [PMID: 15133124]
- Deguchi S, and Sato M. Biomechanical properties of actin stress fibers of non-motile cells. Biorheology 2009; 46(2):93-105. [PMID: 19458413]
- Pellegrin S, and Mellor H. Actin stress fibres. J. Cell. Sci. 2007; 120(Pt 20):3491-9. [PMID: 17928305]
- Adams JC. Formation of stable microspikes containing actin and the 55 kDa actin bundling protein, fascin, is a consequence of cell adhesion to thrombospondin-1: implications for the anti-adhesive activities of thrombospondin-1. J. Cell. Sci. 1995; 108 ( Pt 5):1977-90. [PMID: 7657718]
- Lazarides E. Immunofluorescence studies on the structure of actin filaments in tissue culture cells. J. Histochem. Cytochem. 1975; 23(7):507-28. [PMID: 1095651]
- Heath JP. Behaviour and structure of the leading lamella in moving fibroblasts. I. Occurrence and centripetal movement of arc-shaped microfilament bundles beneath the dorsal cell surface. J. Cell. Sci. 1983; 60:331-54. [PMID: 6348051]
- Heath JP, and Dunn GA. Cell to substratum contacts of chick fibroblasts and their relation to the microfilament system. A correlated interference-reflexion and high-voltage electron-microscope study. J. Cell. Sci. 1978; 29:197-212. [PMID: 564353]
- Small JV, Rottner K, Kaverina I, and Anderson KI. Assembling an actin cytoskeleton for cell attachment and movement. Biochim. Biophys. Acta 1998; 1404(3):271-81. [PMID: 9739149]
- Chen WT. Mechanism of retraction of the trailing edge during fibroblast movement. J. Cell Biol. 1981; 90(1):187-200. [PMID: 7195906]
- Théry M, Pépin A, Dressaire E, Chen Y, and Bornens M. Cell distribution of stress fibres in response to the geometry of the adhesive environment. Cell Motil. Cytoskeleton 2006; 63(6):341-55. [PMID: 16550544]