
How are focal adhesion dynamics regulated?
The highly dynamic and complex molecular composition of focal adhesions is regulated by a number of physico-chemical signals arising from its surroundings, such as extracellular matrix stiffness, topography, and its biochemical composition. The molecular dynamics within focal adhesions, in turn, regulate their functions in sensing substrate rigidity, forming substrate adhesions, cell spreading and migration. In a recent study, Stutchbury et al used fluorescence-based quantitative techniques like FRAP (fluorescence recovery after photobleaching) and FLAP (fluorescence loss after photoactivation) to characterize the mobility of the core components of focal adhesions. They found that the FA core components differed in their mobility that correlated with their functions within the FAs, allowing them to be grouped into distinct functional modules. For instance, proteins connecting the integrins to the actin cytoskeleton were grouped as ‘structural module’ proteins. This module included talin, vinculin, and tensin, all of which showed very low turnover rates. In fact, their turnover was found to depend on the stiffness of the extracellular matrix, suggesting that these proteins primarily function as mechanosensing components. On the other hand, proteins like FAK and paxillin showed high turnover rates and their mobilities were largely unaffected by extracellular matrix stiffness. This implied that FAK and paxillin do not have mechanosensing functions similar to the structural module components. Instead, they were believed to mediate intracellular signaling pathways within the FAs, and were therefore grouped as ‘signaling module’ proteins. Stutchbury et al also noted that mutating key tyrosine residues in FAK and paxillin or treating cells with FAK inhibitor (inhibits FAK autophosphoryaltion) and/or Src kinase inhibitor (inhibits paxillin phosphorylation) led to significant losses in FA functions such as lamellipodia formation, cell spreading and directional cell migration [28302906].
Matrix properties
Substrate rigidity has a great impact on the morphology of certain cell types such as fibroblasts, but has no effect on neutrophils [1]. Rigidity sensing by focal adhesions (FA) can cause changes in FA dynamics and hence propagate signals that alter cell shape and cytoskeletal structures. For example, fibroblast polarization on rigid substrates is preceded by elongation, stabilization and uniform alignment of FAs along the elongation axis of the cell, whereas on compliant surfaces, small rounded and highly dynamic adhesions are formed and no proper polarization is observed [2].
Spacing of matrix ligands is also critical for establishment of stable adhesions. Cavalcanti-Adam et al. have shown that a spacing range of 50-70nm is required between ligand molecules, beyond which adhesions do not form and poor cell migration was observed [3]. This distance seems to coincide with the length of talin, suggesting that talin is the principal mediator of integrin–actin linkage (reviewed in [4][5]). Independent of the global ligand density, it has been shown that nanoclustering of at least four integrin-binding sites in this threshold distance range is essential for effective integrin-mediated signaling [6]. Ligand spacing has also been demonstrated to be important for integrin signaling in endothelial cells [7].
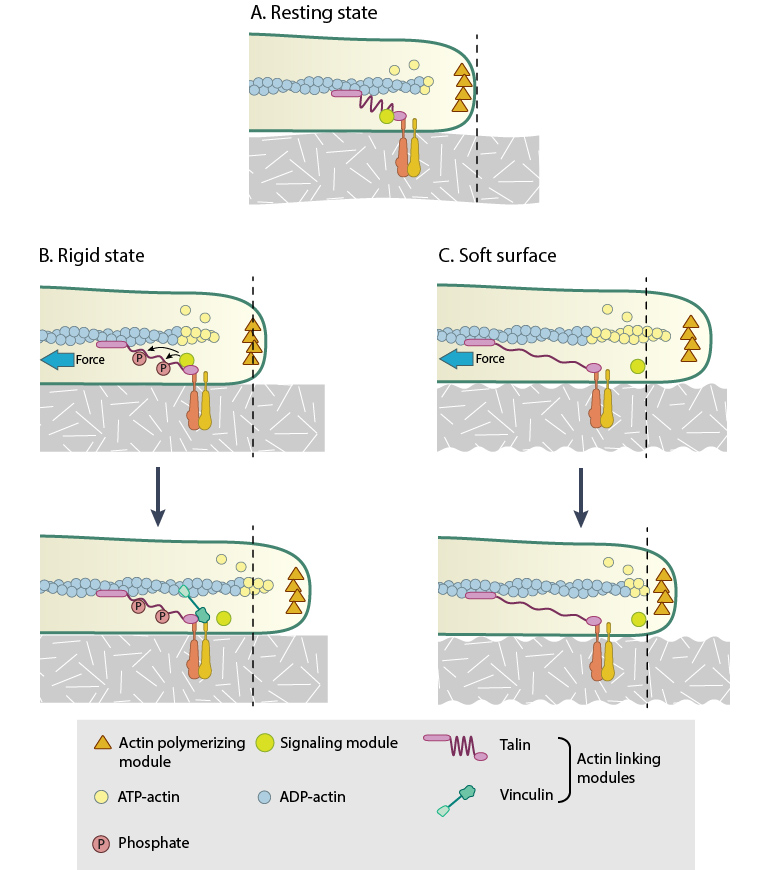
This figure illustrates how force (e.g. substrate rigidity, contraction) hypothetically alters the position of molecules and influences subsequent mechanotransduction events (adapted from [16, 17]). A. In the resting state, actin filaments are linked to the ECM through flexible protein linkers (shown as dark pink ovals with a stretchable coil domain). The essential feature of this model is that the signaling components (e.g. Src-family of protein tyrosine phosphatases [SFKs], shown as a yellow circle) move with respect to the linking proteins, which are moving rearward due to actin rearward movement (represented by theyellow arrow in B and C). B. On rigid ECM substrates, the ECM resists unfolding (shown as a thick blue line on the ECM molecule) due the forces of actin rearward movement. Consequently, a quick rise in force near the cell edge is placed onto the linking proteins (shown by the red coil), which causes unfolding and exposure of sites that can be modified (e.g phosphorylated) by the signaling module components near the adhesion. Modifications to the linking module supports additional protein binding and strengthening of adhesions (shown as a light pink oval). C. On soft ECM substrates, the ECM components are unfolded/stretched due to the forces of actin rearward movement. ECM unfolding correlates to a slower rise in force and delayed unfolding of the linking module components until a point further from the cell edge. Because the signal module components remain at the cell edge, they are displaced relative to the adhesion site and are not available to modify the linking module components to promote reinforcement.
Cycling of integrins is a key aspect of mechanotransduction by focal adhesions. Integrins get activated, cluster, segregate and disassemble throughout the different stages of adhesions. This process facilitates both the transition of early complexes to mature and the ability to rapidly disassemble. All these steps are force-sensitive and dependent on other adhesion components (reviewed in [8]). Recently, it has been proven that integrins rapidly switch between active and inactive conformations within FAs [9]. This influences their trafficking and hence FA dynamics [10].
Adhesion Strengthening
Adhesion strengthening is required for nascent adhesions to survive shear forces exerted by actin retrograde flow. The friction between the adhesion components and hence the strengthening depends on the nature and organization of the receptor-ECM bonds (reviewed in [11][12]), and is known to generally correlate with increasing ligand density [13] and the ability of integrins to cluster laterally [14].
However, the adhesive bonds for β1 integrins are stronger than that of β3 integrins [15][16]. In β1 integrins, the adhesive bond is further enhanced by applied forces that increase the catch bond behavior of the molecule [17], altogether with the engagement of a synergy site in fibronectin [18]. Such catch bond behavior also fits a stochastic model that explains adhesion engagement state in relation to actin retrograde flow and corresponding traction stresses [19]. In β3 integrins, the adhesion strength is increased by linkage to actin [20] and sustained by force-dependent recruitment of components such as talin, vinculin[21][22] and paxillin [23][24], that reinforce the linkage [25] and initiate signaling cascades [26][27]. Adhesion strength can modulate migratory behavior and shape in motile cells [28].
Flow of Actin
Flow of actin (i.e. actin retrograde flow)- The application of mechanical force that is generated by the actin system seems to be a prerequisite for the earliest stages of focal adhesion assembly and generation of traction forces [22][23]. The coupling of the adhesion clutch to moving actin is mediated by the continuous association and dissociation of sliding molecular bonds referred to as ‘stick-slip behavior’ [29][30].Quantification of traction stresses exerted on the ECM by FAs using high-resolution traction force microscopy revealed that intermediate actin flow rates correspond to maximal traction [31]. At the leading edge, where FAs form and flow is high, traction is minimal. However near the larger growing adhesion, the flow is slowed down [31][32] to ~8-10nm/s and is linearly proportional to traction stress [27] (reviewed in [11]).
Two stochastic models explain this biphasic relationship of friction at the moving interface between flowing actin and stationary adhesions [19][30]. The competition between the elastic bonds at the interface and energy dissipation in the viscoelastic actin interior is proposed to modulate the stability of adhesions [30]. The actin-receptor bonds are thought to reach a quasi-equilibrium state between bound and unbound forms. A catch bond model convincingly fits the experimental observations leading to clutch engagement and adhesion growth [19]. These also clarify why traction on the substrate is maximum at intermediate flow [32][33].
Actomyosin Contraction
Myosin contractile forces – FA growth, maturation and dynamics are highly dependent on myosin II contractility, that funnels the internal and applied forces to the adhesions as traction forces ([34], [35], reviewed in [36][37][38], see video below). MyosinIIA has been implicated in adhesion growth, remodeling of actin filaments into stress fibres and their maintenance between adhesions [39][40][41][42] while Myosin-IIB is responsible for the formation of FAs at the rear and hence serve to establish cell polarity [43][44]. However it is to be noted that mature adhesions can withstand about six folds increase in traction stress without any effect on their size [45].
During cell spreading, myosin contraction gets activated at early stages at the end of initial rapid spreading phase, when the cell transforms to a flat morphology, due to the physical barrier posed by the plasma membrane [46]. Thus increase in membrane tension seems to get converted into biochemical signal that activates actin assembly in the protruded area and causes local contraction that favors adhesion assembly [46]. In an independent in vitro study, Yu et al. have shown that upon early actin polymerization from liganded integrin clusters, myosin II contraction of clusters activates protrusion with the actin providing outward forces and the myosin providing the local contraction and inward movement [47]. This cyclic process known as the periodic lamellipodial contraction is mediated by myosin light chain kinase (MLCK) and the forces aid adhesion assembly and complete cell spreading [48]. Hence, one cannot ignore the possibility that the two mechanisms could be interconnected.
References
- Yeung T, Georges PC, Flanagan LA, Marg B, Ortiz M, Funaki M, Zahir N, Ming W, Weaver V, and Janmey PA. Effects of substrate stiffness on cell morphology, cytoskeletal structure, and adhesion. Cell Motil. Cytoskeleton 2005; 60(1):24-34. [PMID: 15573414]
- Prager-Khoutorsky M, Lichtenstein A, Krishnan R, Rajendran K, Mayo A, Kam Z, Geiger B, and Bershadsky AD. Fibroblast polarization is a matrix-rigidity-dependent process controlled by focal adhesion mechanosensing. Nat. Cell Biol. 2011; 13(12):1457-65. [PMID: 22081092]
- Cavalcanti-Adam EA, Volberg T, Micoulet A, Kessler H, Geiger B, and Spatz JP. Cell spreading and focal adhesion dynamics are regulated by spacing of integrin ligands. Biophys. J. 2007; 92(8):2964-74. [PMID: 17277192]
- Giannone G, Mège R, and Thoumine O. Multi-level molecular clutches in motile cell processes. Trends Cell Biol. 2009; 19(9):475-86. [PMID: 19716305]
- Geiger B, Spatz JP, and Bershadsky AD. Environmental sensing through focal adhesions. Nat. Rev. Mol. Cell Biol. 2009; 10(1):21-33. [PMID: 19197329]
- Schvartzman M, Palma M, Sable J, Abramson J, Hu X, Sheetz MP, and Wind SJ. Nanolithographic control of the spatial organization of cellular adhesion receptors at the single-molecule level. Nano Lett. 2011; 11(3):1306-12. [PMID: 21319842]
- Le Saux G, Magenau A, Gunaratnam K, Kilian KA, Böcking T, Gooding JJ, and Gaus K. Spacing of integrin ligands influences signal transduction in endothelial cells. Biophys. J. 2011; 101(4):764-73. [PMID: 21843466]
- Puklin-Faucher E, and Sheetz MP. The mechanical integrin cycle. J. Cell. Sci. 2009; 122(Pt 2):179-86. [PMID: 19118210]
- Rossier O, Octeau V, Sibarita J, Leduc C, Tessier B, Nair D, Gatterdam V, Destaing O, Albigès-Rizo C, Tampé R, Cognet L, Choquet D, Lounis B, and Giannone G. Integrins β1 and β3 exhibit distinct dynamic nanoscale organizations inside focal adhesions. Nat. Cell Biol. 2012; 14(10):1057-67. [PMID: 23023225]
- Arjonen A, Alanko J, Veltel S, and Ivaska J. Distinct recycling of active and inactive β1 integrins. Traffic 2012; 13(4):610-25. [PMID: 22222055]
- Schwarz US, and Gardel ML. United we stand: integrating the actin cytoskeleton and cell-matrix adhesions in cellular mechanotransduction. J. Cell. Sci. 2012; 125(Pt 13):3051-60. [PMID: 22797913]
- Gardel ML, Schneider IC, Aratyn-Schaus Y, and Waterman CM. Mechanical integration of actin and adhesion dynamics in cell migration. Annu. Rev. Cell Dev. Biol. 2010; 26:315-33. [PMID: 19575647]
- Gallant ND, Michael KE, and García AJ. Cell adhesion strengthening: contributions of adhesive area, integrin binding, and focal adhesion assembly. Mol. Biol. Cell 2005; 16(9):4329-40. [PMID: 16000373]
- Selhuber-Unkel C, López-García M, Kessler H, and Spatz JP. Cooperativity in adhesion cluster formation during initial cell adhesion. Biophys. J. 2008; 95(11):5424-31. [PMID: 18689459]
- Petrie TA, Capadona JR, Reyes CD, and García AJ. Integrin specificity and enhanced cellular activities associated with surfaces presenting a recombinant fibronectin fragment compared to RGD supports. Biomaterials 2006; 27(31):5459-70. [PMID: 16846640]
- Roca-Cusachs P, Gauthier NC, Del Rio A, and Sheetz MP. Clustering of alpha(5)beta(1) integrins determines adhesion strength whereas alpha(v)beta(3) and talin enable mechanotransduction. Proc. Natl. Acad. Sci. U.S.A. 2009; 106(38):16245-50. [PMID: 19805288]
- Kong F, García AJ, Mould AP, Humphries MJ, and Zhu C. Demonstration of catch bonds between an integrin and its ligand. J. Cell Biol. 2009; 185(7):1275-84. [PMID: 19564406]
- Friedland JC, Lee MH, and Boettiger D. Mechanically activated integrin switch controls alpha5beta1 function. Science 2009; 323(5914):642-4. [PMID: 19179533]
- Li Y, Bhimalapuram P, and Dinner AR. Model for how retrograde actin flow regulates adhesion traction stresses. J Phys Condens Matter 2010; 22(19):194113. [PMID: 21386439]
- Jiang G, Giannone G, Critchley DR, Fukumoto E, and Sheetz MP. Two-piconewton slip bond between fibronectin and the cytoskeleton depends on talin. Nature 2003; 424(6946):334-7. [PMID: 12867986]
- Humphries JD, Wang P, Streuli C, Geiger B, Humphries MJ, and Ballestrem C. Vinculin controls focal adhesion formation by direct interactions with talin and actin. J. Cell Biol. 2007; 179(5):1043-57. [PMID: 18056416]
- Margadant F, Chew LL, Hu X, Yu H, Bate N, Zhang X, and Sheetz M. Mechanotransduction in vivo by repeated talin stretch-relaxation events depends upon vinculin. PLoS Biol. 2011; 9(12):e1001223. [PMID: 22205879]
- Mofrad MRK, Golji J, Abdul Rahim NA, and Kamm RD. Force-induced unfolding of the focal adhesion targeting domain and the influence of paxillin binding. Mech Chem Biosyst 2004; 1(4):253-65. [PMID: 16783922]
- Zaidel-Bar R, Milo R, Kam Z, and Geiger B. A paxillin tyrosine phosphorylation switch regulates the assembly and form of cell-matrix adhesions. J. Cell. Sci. 2006; 120(Pt 1):137-48. [PMID: 17164291]
- Galbraith CG, Yamada KM, and Sheetz MP. The relationship between force and focal complex development. J. Cell Biol. 2002; 159(4):695-705. [PMID: 12446745]
- von Wichert G, Haimovich B, Feng G, and Sheetz MP. Force-dependent integrin-cytoskeleton linkage formation requires downregulation of focal complex dynamics by Shp2. EMBO J. 2003; 22(19):5023-35. [PMID: 14517241]
- von Wichert G, Jiang G, Kostic A, De Vos K, Sap J, and Sheetz MP. RPTP-alpha acts as a transducer of mechanical force on alphav/beta3-integrin-cytoskeleton linkages. J. Cell Biol. 2003; 161(1):143-53. [PMID: 12682088]
- Barnhart EL, Lee K, Keren K, Mogilner A, and Theriot JA. An adhesion-dependent switch between mechanisms that determine motile cell shape. PLoS Biol. 2011; 9(5):e1001059. [PMID: 21559321]
- Chan CE, and Odde DJ. Traction dynamics of filopodia on compliant substrates. Science 2008; 322(5908):1687-91. [PMID: 19074349]
- Sabass B, and Schwarz US. Modeling cytoskeletal flow over adhesion sites: competition between stochastic bond dynamics and intracellular relaxation. J Phys Condens Matter 2010; 22(19):194112. [PMID: 21386438]
- Gardel ML, Sabass B, Ji L, Danuser G, Schwarz US, and Waterman CM. Traction stress in focal adhesions correlates biphasically with actin retrograde flow speed. J. Cell Biol. 2008; 183(6):999-1005. [PMID: 19075110]
- Alexandrova AY, Arnold K, Schaub S, Vasiliev JM, Meister J, Bershadsky AD, and Verkhovsky AB. Comparative dynamics of retrograde actin flow and focal adhesions: formation of nascent adhesions triggers transition from fast to slow flow. PLoS ONE 2008; 3(9):e3234. [PMID: 18800171]
- Beningo KA, Dembo M, Kaverina I, Small JV, and Wang YL. Nascent focal adhesions are responsible for the generation of strong propulsive forces in migrating fibroblasts. J. Cell Biol. 2001; 153(4):881-8. [PMID: 11352946]
- Cai Y, Biais N, Giannone G, Tanase M, Jiang G, Hofman JM, Wiggins CH, Silberzan P, Buguin A, Ladoux B, and Sheetz MP. Nonmuscle myosin IIA-dependent force inhibits cell spreading and drives F-actin flow. Biophys. J. 2006; 91(10):3907-20. [PMID: 16920834]
- Hotulainen P, and Lappalainen P. Stress fibers are generated by two distinct actin assembly mechanisms in motile cells. J. Cell Biol. 2006; 173(3):383-94. [PMID: 16651381]
- Wolfenson H, Henis YI, Geiger B, and Bershadsky AD. The heel and toe of the cell’s foot: a multifaceted approach for understanding the structure and dynamics of focal adhesions. Cell Motil. Cytoskeleton 2009; 66(11):1017-29. [PMID: 19598236]
- Vicente-Manzanares M, Ma X, Adelstein RS, and Horwitz AR. Non-muscle myosin II takes centre stage in cell adhesion and migration. Nat. Rev. Mol. Cell Biol. 2009; 10(11):778-90. [PMID: 19851336]
- Parsons JT, Horwitz AR, and Schwartz MA. Cell adhesion: integrating cytoskeletal dynamics and cellular tension. Nat. Rev. Mol. Cell Biol. 2010; 11(9):633-43. [PMID: 20729930]
- Even-Ram S, Doyle AD, Conti MA, Matsumoto K, Adelstein RS, and Yamada KM. Myosin IIA regulates cell motility and actomyosin-microtubule crosstalk. Nat. Cell Biol. 2007; 9(3):299-309. [PMID: 17310241]
- Sandquist JC, Swenson KI, Demali KA, Burridge K, and Means AR. Rho kinase differentially regulates phosphorylation of nonmuscle myosin II isoforms A and B during cell rounding and migration. J. Biol. Chem. 2006; 281(47):35873-83. [PMID: 17020881]
- Vicente-Manzanares M, Zareno J, Whitmore L, Choi CK, and Horwitz AF. Regulation of protrusion, adhesion dynamics, and polarity by myosins IIA and IIB in migrating cells. J. Cell Biol. 2007; 176(5):573-80. [PMID: 17312025]
- Rossier OM, Gauthier N, Biais N, Vonnegut W, Fardin M, Avigan P, Heller ER, Mathur A, Ghassemi S, Koeckert MS, Hone JC, and Sheetz MP. Force generated by actomyosin contraction builds bridges between adhesive contacts. EMBO J. 2010; 29(6):1055-68. [PMID: 20150894]
- Lo C, Buxton DB, Chua GCH, Dembo M, Adelstein RS, and Wang Y. Nonmuscle myosin IIb is involved in the guidance of fibroblast migration. Mol. Biol. Cell 2003; 15(3):982-9. [PMID: 14699073]
- Vicente-Manzanares M, Koach MA, Whitmore L, Lamers ML, and Horwitz AF. Segregation and activation of myosin IIB creates a rear in migrating cells. J. Cell Biol. 2008; 183(3):543-54. [PMID: 18955554]
- Stricker J, Aratyn-Schaus Y, Oakes PW, and Gardel ML. Spatiotemporal constraints on the force-dependent growth of focal adhesions. Biophys. J. 2011; 100(12):2883-93. [PMID: 21689521]
- Ji L, Lim J, and Danuser G. Fluctuations of intracellular forces during cell protrusion. Nat. Cell Biol. 2008; 10(12):1393-400. [PMID: 19011623]
- Yu C, Law JBK, Suryana M, Low HY, and Sheetz MP. Early integrin binding to Arg-Gly-Asp peptide activates actin polymerization and contractile movement that stimulates outward translocation. Proc. Natl. Acad. Sci. U.S.A. 2011; 108(51):20585-90. [PMID: 22139375]
- Giannone G, Dubin-Thaler BJ, Döbereiner H, Kieffer N, Bresnick AR, and Sheetz MP. Periodic lamellipodial contractions correlate with rearward actin waves. Cell 2004; 116(3):431-43. [PMID: 15016377]